Philippine lizards of the family Gekkonidae comprise 49 species (Taylor, 1915, 1922; Brown and Alcala, 1978) in 10 genera: Gehyra (1), Gekko (13), Hemidactylus (5), Hemiphyllodactylus (2), Lepidodactylus (6), Luperosaurus (8), Ptychozoon (1), Pseudogekko (4), and Cyrtodactylus (9), (Brown et al., 2007, 2010a, 2011; Welton et al., 2009, 2010a, 2010b; Zug, 2011). An amazing percentage of these species are endemic to the Philippines archipelago (roughly 85%; Brown et al., 2011). Several of the recently described gekkonids in the Philippines were discovered only recently as part of ongoing surveys around the archipelago. Recent phylogenetic studies focused on Philippine gekkonids (Siler et al., 2010; Welton et al., 2010a,b) have resulted in the observation of high levels of genetic diversity among populations of widespread species, an indication that the country's gecko diversity may still be greatly underestimated.
The genus Luperosaurus presently contains 13 species (Brown et al., 2000, 2007, 2010b, 2011). Unfortunately, many of these species are known only from a few specimens (Ota et al., 1996; Brown and Diesmos, 2000). In the Philippines, eight species are recognized: Luperosaurus angliit, Luperosaurus corfieldi, Luperosaurus cumingii, Luperosaurus gulat, Luperosaurus joloensis, Luperosaurus kubli, Luperosaurus macgregori, and Luperosaurus palawanensis (Brown et al., 2007, 2010b, 2011; Gaulke et al., 2007). All of the species in the Philippines, as well as the Borneo species, Luperosaurus yasumai and Luperosaurus sorok (Ota et al., 1996; Das et al., 2008) are robust-bodied (Brown et al., 2011). In contrast, Luperosaurus browni (Peninsular Malaysia and Borneo), Luperosaurus brooksii (Sumatra), and Luperosaurus iskandari (Sulawesi) are recognized to have more slender bodies (Brown et al., 2000, 2011). Brown et al. (2000) conducted the only phylogenetic analysis of this unique radiation of gekkonid lizards to date, and found that the robust- and slender-bodied species are supported to be part of two, reciprocally monophyletic clades, respectively.
Luperosaurus species remain rare in collections, with few exceptions, and little is known of their ecology. Coastal forests are the presumed microhabitat for most species (Brown et al., 2011). Unfortunately, much of this microhabitat has been destroyed across the Philippine archipelago, and many species may currently be threatened due to severe habitat loss.
Luperosaurus corfieldi can be distinguished from congeners by the following combination of characters: (1) medium to large body size (SVL 70.0-95.0 mm); (2) precloaco-femorals 11-19; (3) internasal scales contacting rostral 1–2; (4) scales contacting nostril 4; (5) supralabials 14-16; (6) infralabials 12-14; (7) Toe I scansors 10-14; (8) Toe III scansors 14-20; (9) extent of webbing between Toe III and Toe IV 1/3–1/2; (10) auricular opening subcircular, large; (11) penultimate scansors deeply notched; (12) dorsal tubercles convex; (13) ventrolateral body tubercles few, convex to conical; (14) ventrals slightly enlarged; (15) midbody scales 165; (16) anteriormost chin scales small; (17) anterior forelimb expansions present as wide flaps; (18) posterior forelimb expansions present as wide flaps; (19) anterior hind-limb expansions present as wide flaps; (20) posterior hind-limb expansions present as wide flaps. For extensive comparisons between all Luperosaurus species, see Brown et al., 2011:table 1.
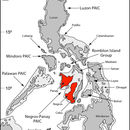
This species is recognized to occur on Negros and Panay islands in the central Philippines.
Visayan (Central) Pleistocene Aggregate Island Complex (PAIC; Brown and Diesmos, 2002).
SVL 70.0-95.0 mm
PNM 7919; type stored in the Philippine National Museum; Philippines, Panay Island, Aklan Province, Municipality of Buruanga, Barangay Tagosip, 11°49.3’N, 121°58.2’E, 450 m elevation.