Dokdonia donghaensis ist ein Bakterium. Der Erstfundort liegt im Meerwasser des Japanischen Ostmeer bei den Liancourt-Felsen.
Die Zellen von Dokdonia donghaensis sind stäbchenförmig, mit einer Breite zwischen 0,3 und 0,6 μm und einer Länge von 1,5 bis 5,0 μm. Flagellen sind nicht vorhanden, auch eine gleitende Bewegung ("gliding motility") wurde nicht beobachtet. Diese Bewegungsart findet man bei vielen Arten der Flavobacteriaceae. Sporen werden nicht gebildet. Die Kolonien sind gelb gefärbt.[1]
Dokdonia donghaensis ist auf Sauerstoff angewiesen, es ist strikt aerob. Der Katalase-Test und der Oxidase-Test fallen positiv aus. Nitrat wird nicht genutzt. Casein wird gespalten, Agar und Stärke werden nicht genutzt. Das dominierende Chinon ist das Menachinon-MK6. Bestes Wachstum erfolgt bei 30 °C und bei 2 % (w/v) NaCl.[1][2] Das Bakterium toleriert relativ hohe Salzkonzentrationen (NaCl) bis zu 7 %, Dokdonia donghaensis ist somit halotolerant.[3] Bei einer Salzkonzentration von 0 % erfolgt kein Wachstum. Die Gram-Färbung verläuft negativ.[1] Das Genom wurde vollständig sequenziert.[4]
Die Arten von Dokdonia sind von Interesse zur Untersuchung der Nutzung von Licht als Energiequelle durch Proteorhodopsine bei Mikroorganismen. Hierzu wurden mehrere verschiedene Arten und noch nicht näher bestimmte Stämme der Gattung untersucht.[5][4] Dokdonia donghaensis nutzt hierbei das Proteorhodopsin (PR) als Protonenpumpe, um mit Licht als Energiequelle ATP zu erzeugen. Dies dient als Ergänzung zu den ansonsten organischen Energiequellen (photoheterotrophes Wachstum). Die Vorsilbe Proteo- des Begriffes Proteorhodopsin bezieht sich auf die Proteobacteria, es tritt nur bei dieser Großgruppe der Bakterien auf. Bei den Archaeen tritt ein ähnliches Protein auf, hierbei spricht man vom Bacteriorhodopsin (BR).
Die mikrobielle Rhodopsine sind lichtaktivierte Pumpproteine. Sie bestehen aus einen einzelnen, von einer Membran umschlossenen Protein, das kovalent an ein lichtempfindliches Pigment gebunden ist.[5] Mit Hilfe der Lichtenergie werden H+ hier Protonen vom inneren der Zelle nach außen transportiert. Hiermit wird ein Konzentrationsunterschied zwischen den Protonen außerhalb (mehr) und innerhalb (weniger Protonen) der Zelle erzeugt. Dieser kann als Energielieferant für die Bildung von ATP durch die ATPase, die die Protonen wieder nach innen transportiert, genutzt werden.
Proterohodopsin ist von vielen marine Bakterien bekannt. Die photische Zone des Ozeans enthält die größte Menge an Bakterien mit PR, durchschnittlich 75 % aller im Plankton vorkommenden Bakterien besitzen die hierfür benötigten Gene.[6] Es kann als eine zusätzliche Energiequelle dienen, wenn Nahrungsmangel herrscht.[7] Eine Vermutung geht davon aus, das der durch das Proteorhodopsin erzeugte Protonengradient (pmf, proton motive force) dazu genutzt wird um Vitamin B1 (Thiamin) aufzunehmen. Das Vitamin B1 ist wichtig für den Kohlenstoffstoffwechsel, einschließlich des Citratzyklus.[6]
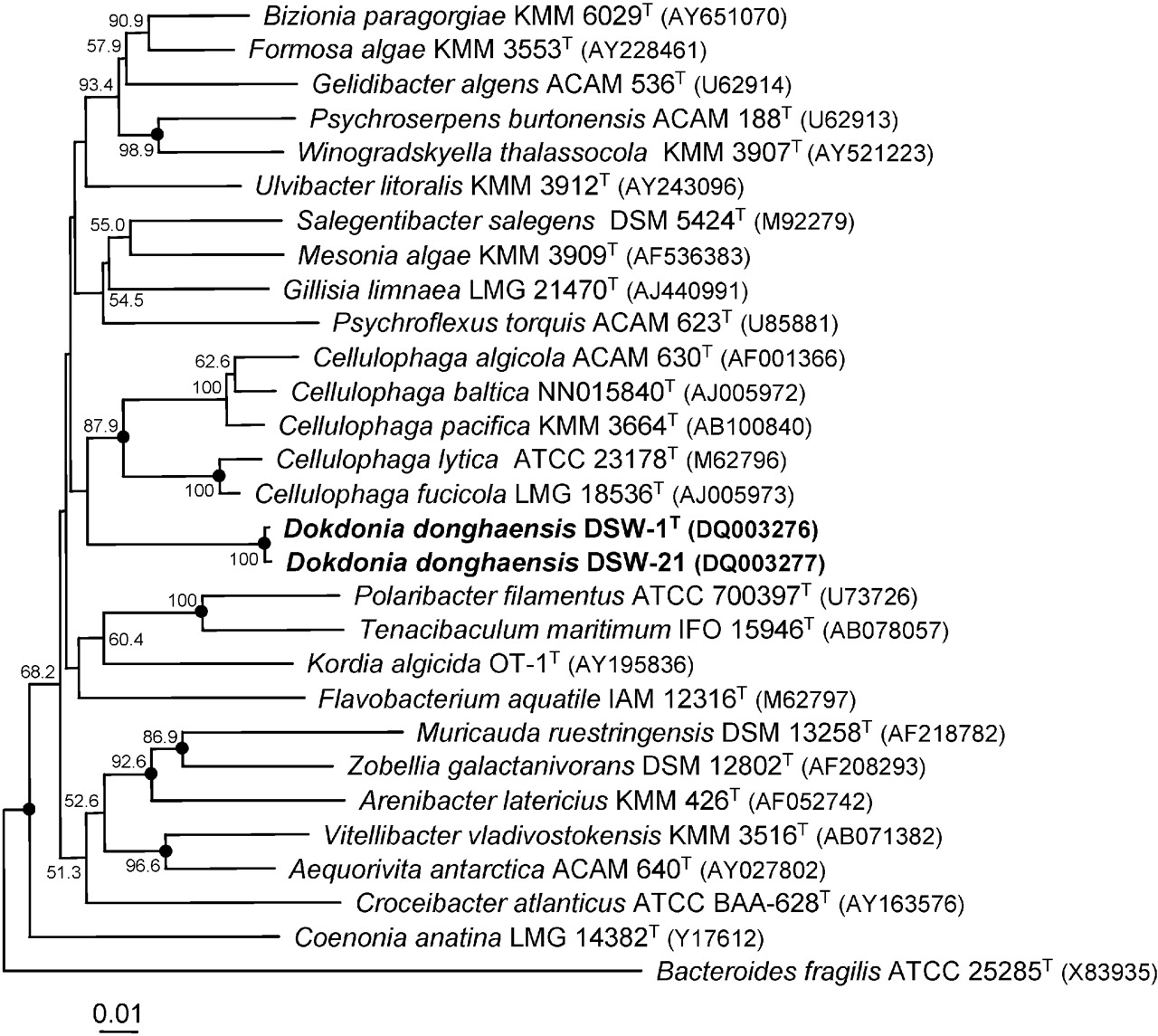
Die Gattung Dokdonia zählt zu der Familie der Flavobacteriaceae. Die Erstbeschreibung stammt aus dem Jahr 2015. Dokdonia donghaensis ist die Typusart, sie ist die zuerst beschriebene Art ihrer Gattung. Im Januar 2022 zählten 10 Arten zu der Gattung.[8]
Dokdonia donghaensis ähnelt der verwandten Gattung Cellulophaga, Unterscheidungsmerkmale sind einmal die fehlende Fähigkeit der gliding motility bei Dogdonia, keine Säurebildung von den getesteten Kohlenhydraten und die Unfähigkeit Agar und Stärke zu spalten.[1]
Dokdonia donghaensis ist ein Bakterium. Der Erstfundort liegt im Meerwasser des Japanischen Ostmeer bei den Liancourt-Felsen.
Dokdonia donghaensis is a strictly aerobic, gram-negative, phototrophic bacterium that thrives in marine environments. The organism can grow at a broad range of temperatures on seawater media. It has the ability to form biofilms, which increases the organism's resistance to antimicrobial agents, such as tetracycline.
Dokdonia donghaensis was discovered and isolated from the Sea of Japan in Korea from an island called Dokdo,[3] a Korean name for the Liancourt Rocks which sovereignty is disputed between Japan and Korea; the date of isolation is not specified. Phylogenetic analysis of the 16S ribosomal RNA gene sequence of the organism revealed two closely related strains, DSW-1T and DSW-21. These strains are classified in Cytophaga-Flavobacterium-Bacteroides (CFB) group, which are dominant genera in marine environments.[4][5] Winogradsky first described Cytophaga-like bacteria as unicellular, gliding, non-spore-formers, and Gram-negative rods, although the shape varies. Many Cytophaga-like bacterial colonies have pigment due to flexirubin-type pigments that are only found in these bacteria and flavobacteria. Cytophaga–Flavobacteria are chemoorganotrophs and are able to degrade biopolymers like chitin and cellulose.[6]
D. donghaensis is a gram-negative bacterium that is rod-shaped, nonmotile, and a non-spore-former. The bacteria are able to grow on seawater media with agar, in which the colonies appear circular, slightly convex, glistening, smooth, yellow, and 1–2 mm in diameter. The DSW-1T strain of D. donghaensis has about 3,923,666 base pairs in its genome.[7] The DNA G+C content for D. donghaensis is 38%.
Image of the phylogenetic tree can be viewed here.
D. donghaensis has not been identified as a human pathogen.
D. donghaensis performs light-driven sodium ion transport. This light-driven proton pump is called a proteorhodopsin, which increases the organism's survival rate because it can convert light into energy that the organism needs to grow.[8] Since the organism uses light for energy, it is a phototroph. D. donghaensis is able to secrete enzymes such as catalase and oxidase.[9] Since the organism is a strict aerobe, it used oxygen as a terminal electron acceptor. The organism can grow on peptone and tryptone as the sole carbon and nitrogen sources. D. donghaensis cannot grow in the absence of NaCl or when it is greater than 7% (w/v); growth is optimal at 2% (w/v) NaCl. The organism is susceptible to the antibiotics tetracycline and carbenicillin,[10] although the organism's ability to form biofilms makes it difficult for antibiotics to penetrate the viscous layers.
D. donghaensis lives in halophilic marine environments. The organism can grow as low as 4 °C and as high as 35 °C, although the optimum temperature for growth is 30 °C. The optimal pH for growth is 7–8, although growth can be observed at a pH as low as 5.5.[11]
D. donghaensis is able to form biofilms in marine habitats, which is a survival strategy that allows the organism to grow while being protected from environmental stresses. Biofilm formation serves a purpose for marine bacteria in that it increases their resistance to antimicrobial agents, desiccation, and grazing.[12][13][14][15] Biofilms allow the microbes to attach to surfaces by excreting extracellular polymeric substances (EPS). Marine bacteria that adhere to surfaces form host-specific and spatially structured communities that are fairly stable.[16][17][18]
Dokdonia donghaensis is a strictly aerobic, gram-negative, phototrophic bacterium that thrives in marine environments. The organism can grow at a broad range of temperatures on seawater media. It has the ability to form biofilms, which increases the organism's resistance to antimicrobial agents, such as tetracycline.
ドクドニア・ドンヘンシス(Dokdonia donghaensis)は、海洋環境を好む絶対好気性でグラム陰性の光栄養細菌の一種である(→形態とゲノム)。海水培地中で生育可能な温度幅は広い(→生育環境)。バイオフィルムを形成する能力を持つ。当該バイオフィルムは、テトラサイクリンなどの抗生物質に対する本バクテリアの耐性を向上させる(→代謝)。
Yoon et al. (2005) は「東海にある韓国領の、独島と呼ばれる島から」、Dokdonia donghaensisを発見し、分離したと報告した[1]。分離の日付は特定されていない。本細菌の16SリボソームRNAの塩基配列に基づく系統解析によると、DSW-1T と DSW-21 の2系統と近い関係にあることがわかった。この2つの系統は、Cytophaga類縁細菌群(Cytophaga-Flavobacterium-Bacteroides (CFB) group)に分類され、海洋環境において優勢な属である[2][3]。セルゲイ・ヴィノグラツキイは、Cytophaga類縁細菌群が、形態こそ多様であるものの、単細胞生物であり、滑走(英語版)を行い、芽胞を作らず、グラム陰性桿菌であることが共通していることを細菌研究史における早期に記述している。多くのCytophaga類縁細菌群の菌叢は、同細菌群とフラヴォバクテリア(英語版)にしか見られないフレキシルビン(英語版)型色素に由来する生物学的色素(英語版)を持つ。Cytophaga類縁細菌群は化学合成有機栄養生物であり、キチン質やセルロースのような生体高分子(バイオポリマー)を分解できる[4]。
D. donghaensis は好塩性であり海洋環境に生息する。生息可能温度は 4 °C から 35 °C まで。生長に適した温度は 30 °C。生長に適した pH は 7-8 であるが 5.5 より低い pH でも生長が観察された[5]。
Kim et al. (2016) による「東海(韓国海)中にある独島沿岸の海水から分離された、プロテオロドプシンを含む海洋性のフラヴォバクテリウム・ドクドニア・ドンヘンシス DSW-1T 株の完全なゲノム配列」と題した論文によると、D. donghaensis は、桿状の外観を呈し、非運動性で、芽胞を作らないグラム陰性菌の一種である[6]。海水寒天培地上で培養させることが可能であり、その菌叢は直径 1-2 mm の円形を呈する[6]。また、わずかに盛り上がり、ツルツルしており、光を反射して黄色に輝く[6]。D. donghaensisの DSW-1T 菌株は、その遺伝情報全体(ゲノム)の中に約3,923,666の塩基対を有する[6]。D. donghaensis のデオキシリボ核酸のグアニン+シトシン含量は、38% である[6]。
Microbiology Society が系統樹を提供している。IJSEM への登録は、「独島東海」と名づけられた本種のほか、「独島韓国(Dokdonella koreensis)」、「ヴァージバシラス独島(Virgibacillus dokdonensis)」、「マリバクター独島(Maribacter dokdonensis)」、「マリノモナス独島(Marinomonas dokdonensis)」と同時に行われた[7]。このような韓国籍人による新種の微生物の命名の動向については、何らかの地政学的目的があると推定されている[8]。
D. donghaensis は光によって駆動されるナトリウムイオン輸送を行う。この光駆動性プロトンポンプは、プロテオロドプシン(英語版)と呼ばれている。プロテオロドプシンは光を、微生物の生長に必要とするエネルギーへと変換できる。そのため、微生物の生存率を増加させる[9]。D. donghaensis は光を栄養にするので、光栄養生物である。D. donghaensis は、カタラーゼやオキシダーゼといった酵素を分泌することができる[10]。D. donghaensis は偏性好気菌(strict aerobe)であるため、酸素を最終電子受容体として用いる。D. donghaensis は、ペプトンとトリプトン培地を唯一の炭素・窒素源として生育可能である。D. donghaensis は、塩化ナトリウムがまったくないか、あるいは溶液濃度で 7% (w/v)を越えた環境下では生育不能である。生長には 2% (w/v) NaCl であることが好ましい。D. donghaensis は、抗生物質であるテトラサイクリン(テトラサイクリン系抗生物質)とカルベニシリンに弱いが、抗生物質が透過しにくい粘性のあるバイオフィルムを形成する能力を有する[11]。
D. donghaensis は、海洋環境においてバイオフィルムを形成することができる。これは環境負荷から保護された状態で生長を図るという生存戦略の一つである。海洋細菌がバイオフィルムを形成する目的は、抗菌因子、乾燥、接触による細胞の損傷に対する抵抗力を増加させることにある[12][13][14][15]。バイオフィルムにより、微生物は細胞外高分子物質(EPS: Extracellular polymeric substances)を排泄することで海面に居続けることができる。海面に固着する海洋細菌は、宿主特異的で空間的に構築された、安定性の良好な微生物群落を形成する[16][17][18]。
2017年1月時点において、D. donghaensis がヒト病原体(英語版)であるか否かは確認されていない[要出典]。
{kind=link}