
This description covers characteristics that may be relevant to fire ecology and is not meant for identification. Keys for identification are available (e.g., [17,19,36,54]).
At maturity, marsh thistle is generally an erect forb with a single branching stem. Plants may reach 6 feet (2 m) when flowering [17]. Stems are spiny and winged because a portion of the leaf blade is attached to the stem. Most plant parts are covered with long sticky hairs [37,54].
Photo © J.C. Schou; Biopix.dkMarsh thistle is best described as a monocarpic perennial. Plants often reproduce within 2 years but may take longer [19,36,56]. Rosette leaves are long, spiny, and deeply lobed [17,83]. In Jutland, Denmark, many marsh thistle plants remained vegetative for 2 or more years. All plants died after flowering, unless they were damaged during the flowering stage, in which case they regrew the next year [4]. Another researcher reported that marsh thistle generally flowered at 4 years old in its native range (Linkola 1935 cited in [6]). In 2 seashore meadows in Tullgarn, Sweden, most (68-86%) marsh thistles flowered 3 years after germination, while the rest flowered later. In this area, the researcher noted a small number of marsh thistle plants flowering twice [48]. Damage to these plants was not reported, but grazing and trampling were common in the area. (See Vegetative regeneration for another report of marsh thistle flowering in successive years.)
On flowering stems, alternate leaves are 6 to 12 inches (15-30 cm) long near the base but shorter near the top. Leaf spacing is generally much wider near the top than at the base of the plant [17,83]. Spines up to 6 mm long occur along leaf margin lobes [17].Marsh thistle flower heads also appear spiny [83]. Perfect disk florets occur in heads that develop on short branches at the end of the stem [19,36, 36,79]. Few to many flower heads may be produced [17]. Heads typically measure 0.4 to 0.6 inch (1-1.5 cm) across [79].
Marsh thistle produces achenes that measure between 2.5 and 3.5 mm and are attached to a feathery pappus of fine cottony hairs [17,19,37, 83]. The pappus is typically less than 0.4 inch (1 cm) long [37], and achenes average 2 mg (Grime and others 1988 cited in [28]).
Marsh thistle produces a taproot with clusters of fibrous roots [17, 36]. It lacks rhizomes [54]. In Jutland, Denmark, root development of vegetative marsh thistles was described in detail. Increased root growth coincided with increased leaf growth and increased flowering probability. Twenty-three percent of plants in the 3rd vegetative life stage (described in the table below) flowered the next year, while 79% or more plants in 4th vegetative life stage flowered the next year. All plants died after flowering [4].
Photos ©Steve Garske, Great Lakes Indian Fish & Wildlife CommissionSimilar native species: In the Great Lakes, New England, and eastern Canada, marsh thistle occupies habitat similar to that of the native swamp thistle (Cirsium muticum). Swamp thistle lacks stem spines and produces less spiny-looking flower heads than marsh thistle [83].
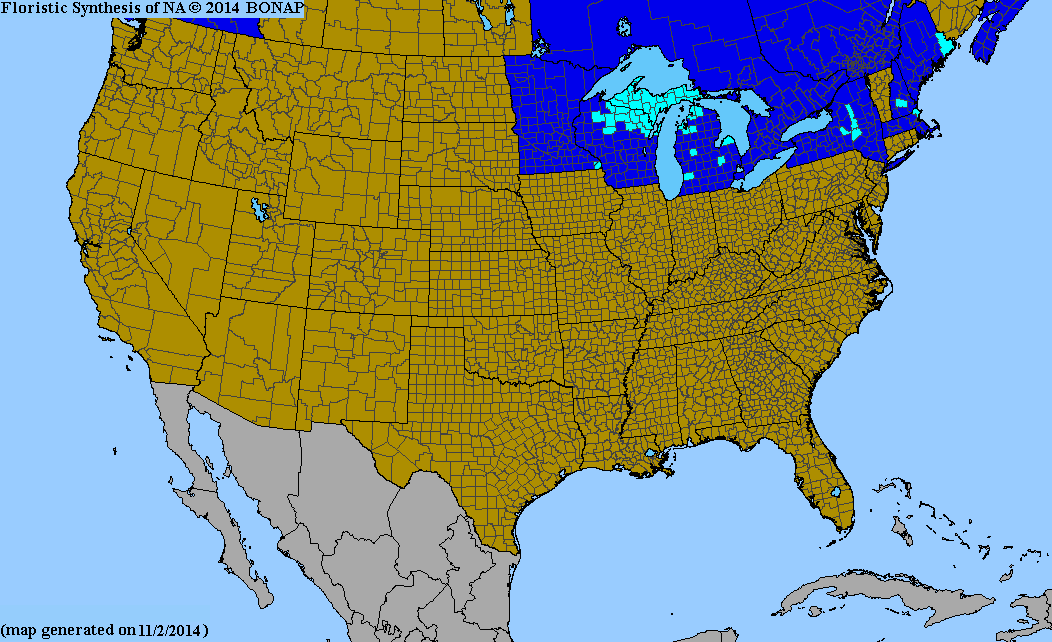
Marsh thistle occurs as a nonnative species in Wisconsin, Michigan, New York, Massachusetts, and New Hampshire in the United States and north to Newfoundland, Nova Scotia, Quebec, Ontario, and British Columbia in Canada [37,56,69]. It is native to Europe [17] and was first reported in North America in the early 1900s [79]. In the Great Lakes area, marsh thistle populations are considered "vastly under reported", although marsh thistle was reported in 10 northeastern counties in Wisconsin as of 2007 and is considered well established in Michigan. Populations in Canadian provinces and northeastern US states are scattered [56]. Plants Database provides a distributional map of marsh thistle.
Marsh thistle was reported in New England by 1902 and in the Great Lakes area by 1934 (review by [56]). In New Hampshire, marsh thistle was first reported from East Andover in 1902. Plants occurred over a nearly 20-acre (8 ha) area in a "moist forest tangle" that was more than a mile (1.6 km) from the nearest town or cultivated area. Method of introduction was unknown. Marsh thistle was reported in South Boston in 1908 and in Newfoundland in 1910 [37]. In 1944, marsh thistle was reported in several communities near Halifax, Nova Scotia [54]. In Michigan, marsh thistle was first collected from Marquette County in 1934 [79]. It was first recorded in Wisconsin in 1961 [56]. In British Columbia, marsh thistle was first reported in 1954 [41]. Marsh thistle was reported during a 1964 survey of a ponderosa pine (Pinus ponderosa) forest in northwestern Nebraska [40]. However, no other sources reported marsh thistle in Nebraska as of 2009, suggesting that this population was transient or incorrectly identified.
Local distribution changes: The range of marsh thistle in North America is "rapidly expanding". Marsh thistle populations in Europe occur almost as far north as the Arctic Circle, suggesting marsh thistle could grow and spread throughout the boreal forest regions of North America [17]. In Wisconsin and Michigan, the area occupied by and the abundance of marsh thistle have increased since its introduction and continue to increase [56]. Marsh thistle was first recorded in Michigan in 1934 and by 1956 was considered locally frequent and common in Michigan's Upper Peninsula and on islands in the Straits of Mackinac [78,79]. By 1959, marsh thistle spread to the Lower Peninsula, and it continues to spread south. "Dense, ungainly colonies" occupy miles of ditch banks in Michigan, and populations have spread into adjacent natural areas [79]. Although marsh thistle has occurred in British Columbia since the 1950s, its spread has been more recent. A "diminutive patch" of marsh thistle west of McBride, British Columbia, was reported in 1991, but marsh thistle had spread at least 130 miles (210 km) by early 2000. Spread occurred primarily along roadways and through river valleys [41]. For more on the potential impacts of marsh thistle persistence and spread, see Impacts and Control.
Fuels: Marsh thistle fuel characteristics and influences on native fuel beds were not described in the available literature (2009).
FIRE REGIMES: Marsh thistle is most common in moist to wet habitats (see Site Characteristics), where fires may be rare and/or burn with low severity. However, on sites with deep organic soils, fires may be infrequent but severe. Altered FIRE REGIMES in areas invaded by marsh thistle habitats were not reported. Fire studies in sites invaded by marsh thistle are needed.
See the Fire Regime Table for further information on FIRE REGIMES of vegetation communities in which marsh thistle may occur. Find further fire regime information for the plant communities in which this species may occur by entering the species name in the FEIS home page under "Find FIRE REGIMES".
Marsh thistle seeds generally germinate best in warm temperatures and full light after cold stratification. However, some germination occurs without stratification, in cool temperatures, and in the dark. Pollination method and parent plant litter can also affect germination.
During field studies in Tullgarn, Sweden, germination percentages for marsh thistle were very low; 0.2% to 0.4% germination in one meadow population and 9% to 17% germination in another meadow population [48].
Temperature, light, and moisture effects: Cold stratification and high light and moisture levels may produce the highest germination percentages for marsh thistle seeds, but seeds may also germinate without cold stratification and in the dark. Warm temperatures (≥54 °F (12° C)) are typically best for germination regardless of prior chilling and light conditions [24,45,75].
Increasingly cold temperatures significantly (P<0.001) increased the germination of marsh thistle seeds collected from a wet meadow in the Czech Republic. Germination was highest but was still less than 40% after 30 days at 6.8 °F (-14° C). At 68 °F (20° C), germination was less than 20% [32]. Marsh thistle seeds collected from plants in the Netherlands, however, "gave no problems in germination", although other species required winter temperatures before germinating [43]. Marsh thistle seeds collected in August from a sphagnum bog in England's Sheffield area also germinated well (91%) without cold stratification. After 3, 6, and 12 months of storage at 41 °F (5° C), germination was 50%, 88%, and 79%, respectively. In full light, germination was 90%; in the shade (2.4% of full light), germination was 86%. In the dark, germination was 36% [24]. In other laboratory studies, germination of freshly collected marsh thistle seed ranged from 32% to 72% in the light and 0.8% to 20.7% in the dark. Storage at 39 °F (4 °C) reduced germination in the light and dark [75].
Laboratory findings suggested that marsh thistle seeds could germinate beneath ash canopies, but seedlings were rare in the understory. At low red/far red (R/FR) light levels that were slightly lower than those penetrating ash thickets in the Netherlands, freshly harvested or dry-stored marsh thistle seed germination was lower than at high R/FR light levels. Cold stratification increased germination at low R/FR light levels and at low temperatures. Germination failed even with stratification at a R/FR level of less than 0.2 [45].
Marsh thistle seeds failed to germinate at water stress levels of 0.25 MPa in the laboratory. Germination ranged from 69% to 57% up to water stress levels of 0.1 MPa but was reduced to 3.8% at 0.2 MPa [75].
Timing and parent plant effects: Researchers found that in a spring area in central Jutland, Denmark, marsh thistle seedlings from fall-germinating seeds were generally larger than those from spring-germinating seeds [4]. Seedlings were rare beneath flowering marsh thistles or in the immediate vicinity of marsh thistle rosettes. In the laboratory, germination of marsh thistle seeds was significantly (P<0.05) lower when treated with extracts of marsh thistle leaf material than when untreated [3]. Seedling growth may also be reduced in soil with marsh thistle leaf litter (see Seedling establishment, growth, and survival).
Pollination method effects: In the Netherlands, marsh thistle seeds from plants protected from insects germinated at a greater percentage and rate than seeds from cross-pollinated plants. After 4 days, germination of seeds from cross-pollinated plants (1.6%) was significantly (P<0.01) less than that of seeds from protected plants (19.7%). After 14 days, germination differences were still significant (P<0.05); 77.4% of seeds from cross-pollinated plants and 87.7% of seeds from protected plants germinated [73]. Seedling emergence and survival may also be reduced in cross-pollinated plants (see Seedling establishment, growth, and survival).
Throughout its range, marsh thistle is common in disturbed habitats including roadsides, fields, yards, gardens, and early-seral forests or clearings [36,37,83]. In New England, marsh thistle is considered possible in any moist disturbed area [37]. Marsh thistle also occurs in less disturbed habitats including thickets, damp woods, forest edges, and wetlands [36,37,83].
Climate: In North America, marsh thistle is most common in moist areas with long cold winters [56]. During a survey of major roadways in South Island, New Zealand, marsh thistle was generally restricted to cool, wet areas in a zone where the annual water deficit was less than 2 inches (50 mm) [81].
Climates are described from several European marsh thistle habitats, which may allow for a better prediction of its spread potential in North America. In Europe, marsh thistle populations occur almost as far north as the Arctic Circle, suggesting populations in North America could potentially spread through the boreal forest zone [17]. In southeastern Scotland, marsh thistle is common in grasslands where the average February and July temperatures are 34.2° F (1.2° C) and 57° F (13.9 °C), respectively, and annual precipitation averages 35.2 inches (894 mm) [31]. In south Wales, the climate is mild and oceanic. Temperatures average 39° F (4° C) in February and 59° F (15° C) in July [26]. In the central and northeastern Netherlands, marsh thistle is common in sedge fens where annual precipitation averages 30.1 to 31.7 inches (765-806 mm) [7]. In wet meadow marsh thistle habitats in Ceske Budejovice, Czech Republic, annual precipitation averages 24.4 inches (620 mm), and minimum and maximum temperature averages for July are 52.9° F (11.6° C) and 75.4° F (24.1° C) and for January are 20.8° F (-6.2° C) and 33.1° F (0.6° C) [32].
Elevation: Marsh thistle occurs at elevations from 30 to 2,600 feet (10-800 m) in North America [17].
Soils: In marsh thistle habitats in North America, soils were rarely described in detail. In Canada, marsh thistle grew in organic wetland soils and in coarse gravelly soils along roadsides. Large populations and high densities were often associated with high water tables [41]. Marsh thistle is common in moist, acidic soils in New England [37] and Wisconsin [83].
In Europe, studies of soils in marsh thistle habitats suggest a wider tolerance of soil textures and pH levels than those evident from the few North American studies and sources available as of 2009. In southeastern Scotland, marsh thistle was most common on poorly to very poorly drained, acidic (pH 5-5.5), clay soils [31]. In southwestern England, marsh thistle seedlings emerged from basic soil samples collected from a 45-year-old oak woodland but not from acidic soils collected from the same woodland [80]. However, this study does not necessarily imply an establishment preference for basic soils and could simply be a result of uneven seed dispersal. In an old field in the Geescroft Wilderness area of England, marsh thistle occurred in damp, cool, acidic heavy loams but not in a field described as wet and alkaline [8]. This finding could also be the result of dispersal and not preference. In the western part of the Utrecht Province in the Netherlands, marsh thistle was significantly (P<0.01) more frequent on ditch banks adjacent to fields fertilized with low levels of nitrogen (0-250 kg N/ha/year) than on fields fertilized with high levels of nitrogen (250-500 kg N/ha/year) [77]. Marsh thistle emerged from all organic peat soil samples taken from a "recently" clearcut birch woodland in Germany. Soils were fed with calcium-rich groundwater [5].
Moist conditions are typical in marsh thistle's native habitats, but plants may not tolerate long-term flooding or saturation. In the coastal dune areas of the Netherlands, marsh thistle is common in oneseed hawthorn-European white birch woodlands where ground water occurs at or near the soil surface [70]. In a spring area of Jutland, Denmark, marsh thistle plants did not grow in the wettest areas or on "regularly flushed" springs [4]. Marsh thistle's drought tolerance is likely low. After a 3-month drought, leaves from marsh thistle plants growing in a species-rich, calcareous grassland in Derbyshire, England, had low relative water content. Of leaves of the 31 plant species evaluated, marsh thistle leaves were ranked 6th lowest in relative water content. Monthly precipitation during the drought averaged 43%, 84%, and 26% of long-term monthly averages [11].
European habitats and plant communities:
In Europe, marsh thistle occurs in fens, sedge (Carex spp.)
meadows, and moist grasslands and woodlands. In Suffolk, England, the bluntflower rush (Juncus subnodulosus)-marsh thistle fen occurred where shrub vegetation was cleared [18]. Marsh thistle also occurs in the understory of mires
at Holmsley Bog in Hampshire, England. Overstory species, which may reach 12 feet (3.7 m) tall, often include English oak (Quercus robur) and/or English yew (Taxus baccata) [20]. In the British Plant Communities books, marsh thistle is reported in:
In southeastern Scotland, marsh thistle is common in colonial bentgrass (Agrostis tenuis) grasslands on poorly drained sites [31]. On coastal dunes in the Netherlands, marsh thistle is common in oneseed hawthorn-European white birch (Crataegus monogyna-Betula pendula) woodlands with ground water at or near the soil surface [70]. Marsh thistle is also described in surface water-fed sedge fens [7], bare sandy sites, and open sites with chee reedgrass (Calamagrostis epigejos) and seaberry (Hippophae rhamnoides) [73].
Marsh thistle rosette diameter and probability of flowering are positively correlated. Rosette diameter and probability of flowering generally decrease as densities or canopy cover of associated vegetation increase.
In a greenhouse study, marsh thistle flowering was positively correlated with rosette size (r =0.40, P<0.05), and rosette size was negatively correlated with marsh thistle plant density (r =-0.38, P<0.05). The percentage of flowering plants was greatest when plant density was lowest and rosette diameter was greatest. Marsh thistle plants at the lowest density produced significantly (P<0.05) more fruits than those at moderate and high densities [15].
Growth and reproductive fate of marsh thistle plants grown in different densities in the greenhouse [15] Density (plants/0.25 m²) 1 2 4 Average rosette diameter (cm) 34.8* 25.6* 20.6 Percentage of individuals flowering in 2nd year 90 60 25 Height of flowering shoot (cm) 139.7* 116.9* 102.2 *Values within the row are significantly different (P<0.05).In field studies in the Reski Range of Poland's Bialowieza Primeval Forest, marsh thistle population dynamics were studied within a single cohort and for many cohorts within different vegetation types. In a grassland area, a spring-emerging marsh thistle cohort was studied for 5 years. No plants flowered in their 1st year, and only 30% flowered in their 2nd year. The highest percentage of plants flowered in their 4th year [15].
Fate of a marsh thistle seedling cohort monitored over 5 years in a grassland in Poland [15] Plant age (years) Survival (%) Flowering (%) Rosette diameter (cm)As succession proceeded to grass-, forb-, patchy sedge-, and willow-dominated vegetation (5, 10, and 15 years after mowing) in the Bialowieza Primeval Forest, Falinska [15] conducted many studies and made several observations on marsh thistle populations, concluding that:
Marsh thistle flowers are self-compatible, but most are cross-pollinated by insects. In meadows in Tullgarn, Sweden, marsh thistle flowers were primarily pollinated by bumblebees [48]. In mountain and sea cliff habitats in England, bees were the dominant daytime pollinators [39].
Seeds from self-pollinated plants may have lower viability than seeds from cross-pollinated plants. Self-pollinated marsh thistle plants from mountain habitats in Monmouthshire and sea cliff habitats in Gower, England, produced significantly less viable seed than cross-pollinated plants [39]. Field experiments in 3 marsh thistle populations north of The Hague in the Netherlands revealed no significant differences in the germination of seed from cross-pollinated plants and plants protected from insects. Marsh thistle populations occurred in a dense birch (Betula spp.) woodland, a grassland, and a bare sandy site. Seeds produced by cross-pollinated plants weighed significantly (P<0.01) less than those produced by protected plants. Although some dispersal had occurred by the time production was evaluated, seed production of cross-pollinated plants exceeded that of protected plants by as much as 58.9%. Production differences were not significant [73]. Germination, seedling establishment, and plant survival based on pollination method are discussed more in the sections below.
Although many have studied marsh thistle seed bank dynamics and attempted to determine the longevity and persistence of seed in the soil, findings and conclusions from these studies disagree. Some suggest a short-lived seed bank [50], while others suggest a persistent seed bank [46,63]. Methodology and scope of marsh thistle seed bank studies differ, making them difficult to compare and evaluate. It is possible that a portion of marsh thistle seed germinates immediately following dispersal [50], but a smaller portion fails to germinate, becomes dormant, and develops germination requirements different from those of fresh seed [46]. Of seeds collected from plants near The Hague, 40% germinated after 1 year of burial [76].
Experiments conducted in Wellesbourne, England, suggested that seeds did not persist more than 3 years in the soil. Marsh thistle seeds were mixed with soil and put in a container that was sunk into the ground in September. Soil was mixed 3 times/year and emergence monitored. Most seeds germinated within a year and most germinated in the spring, although some fall germination occurred. Not all sown seeds were recovered, and the researcher suspected that seeds covered by 3 inches (7.5 cm) of soil germinated but failed to emerge [50].
Emergence of marsh thistle seeds over time in England [50] Time in the ground 4 months 1 year 2 years 3 years 4 years 5 years Percentage of seeds emerging 8.8% 33.3% 0.2% 0.1% 0 0Increasing depth of burial increased the survival of buried marsh thistle seed in the Netherlands. In a field experiment, 4% of marsh thistle seeds survived 27 months of burial at 2- to 5-inch (5-10 cm) depths, and 40% of seeds survived the same amount of time at 6- to 8-inch (15-20 cm) depths [75].
Soil samples collected from 95- to 150-year-old European beech (Fagus sylvatica) woodlands in southern Sweden rarely contained marsh thistle seed. Soil samples were collected in April at least 330 feet (100 m) inside the woodland edge. Surveys revealed no marsh thistle seedlings in the woodland. Fourteen marsh thistle seedlings/m² emerged from the top 2 inches (5 cm) of mineral soil collected from 1 of the 7 sampled sites. Researchers noted that after clearcutting, marsh thistle was common in the area. Because marsh thistle did not emerge from all soil samples, researchers supposed that emergence on cleared sites resulted from recent long-distance seed dispersal and not a persistent seed bank [60].
Based on several field and greenhouse observations and experiments, Pons [46] concluded that marsh thistle seed does persist in the soil seed bank. In the Netherlands, marsh thistle is common following cutting in ash (Fraxinus spp.) stands. Marsh thistle seedling emergence was compared in soil samples taken from sites cut 7 years earlier. Soil was collected from an undisturbed site and a site where soil had been artificially disturbed. Just 19 marsh thistle seedlings/m² emerged from undisturbed soils, and 497 marsh thistle seedlings/m² emerged from disturbed soils, suggesting that emergence was not limited to wind-dispersed seed on the soil surface but also came from soil-stored seed that was encouraged to germinate by the soil disturbance. In multiple follow-up experiments, Pons concluded that dormancy in marsh thistle seeds was triggered by high temperatures and reduced light levels. Exposure to light was the principal stimulus for germination of soil-stored seed, and brief light exposure during winter harvesting could allow for emergence several months later [46].
In a meadow in Poland's Bialowieza Primeval Forest, the density of marsh thistle in the soil and in aboveground vegetation generally decreased as succession progressed. The meadow was managed with regular mowing that was discontinued when the study began. Marsh thistle plants and seeds occurred in all stages of succession, 0 to 20 years since the last mowing, but density generally decreased as time since last mowing increased [16]. Marsh thistle survival, growth, and development were also studied as succession progressed in this meadow (see Plant development and survival).
Density of marsh thistle seed in the soil from a meadow as time since last mowing increased [16] Time since last mowing (years) 0 5 10 15 20 Dominant vegetation Grasses Grasses, increased proportion of tall forbs Forbs Sedges, some willow clumps Willow clumps, still some tall herbs and sedges Marsh thistle seed bank density (seed/m²)* 320 393 217 144 50 *Determined by separating seeds from soil; 40 samples (10×10×3 cm) were collected at each 5-year interval.
Wind is likely the most common dispersal mechanism for marsh thistle seeds, but seeds may also be dispersed by gravity, in water runoff, or by animals and equipment [41,42]. Marsh thistle seeds are attached to a "thistle-down" pappus that aids in wind dispersal [83]. A review reports that while most marsh thistle seeds fall within 33 feet (10 m) of the parent plant, high winds may carry seeds several kilometers [41]. Wind dispersal distances may be reduced by increased densities and heights of neighboring plants [57], decreased heights of marsh thistle plants [74], and increased seed weights [73].
From laboratory experiments and achene and pappus measurements, researchers calculated that marsh thistle seeds could be dispersed a maximum of 22 feet (6.8 m) in 10 mile (16.4 km)/hour winds. Seeds released from a 36-inch (90 cm) height traveled a maximum of 7.5 feet (2.3 m) and 15 feet (4.5 m) in wind speeds of 3.4 miles (5.5 km)/hour and 6.8 miles (10.9 km)/hour, respectively. Researchers noted that dispersal distances would likely be less in dense stands, where wind speeds are reduced and obstructions are increased [57]. Based on these calculations, researchers suggested seed dispersal distances could be reduced 35% to 50% when marsh thistle plant heights were reduced 16% by larval insects feeding on plant stems [74]. Dispersal distance changes as a result of the greater seed weights of self-pollinated plants were not calculated, but researchers suggested that heavier seeds fell closer to the parent plants [73].
During seed trapping studies conducted in peat-harvested areas in Finland, marsh thistle seeds may have dispersed distances of 160 feet (50 m) or more. In one area, 1 marsh thistle seed/m² was recovered from seed traps located 820 feet (250 m) from the forest edge. There were no mature marsh thistle plants reported in the trapping area. Seed traps 160 feet (50 m) from the forest edge collected 16 marsh thistle seeds/m² [55]. This study, however, was not designed to directly estimate seed dispersal distances. In another seed-trapping study in an eastern Scotland grassland, marsh thistle was collected from traps but not from soil in plots where vegetation was herbicide-killed. Traps contained less than 10 marsh thistle seeds/m², and the distance to the nearest mature plants was not reported. Researchers indicated that seeds were likely transported by wind, but may have been transported in surface water runoff or in animal fur or feathers [42].
It is likely that marsh thistle seeds are transported by equipment, but direct evidence is lacking. A review suggests that logging equipment may have transported marsh thistle seed. In British Columbia, new marsh thistle populations have been reported on mechanically-disturbed sites hundreds of kilometers from existing populations [41]. In hay fields in the northern Netherlands, marsh thistle was present but its seed was not collected from haying equipment used for mowing in August [62]. Although seeds were not recovered from mowing machinery, haying equipment may still have contributed to marsh thistle seed dispersal. It is possible that seeds had fallen off before counts were made or that seed was dispersed in the mowed area by blowing motors.
Reviews report that a single marsh thistle plant may produce up to 2,000 seeds [41,56]. A vernalization period is considered necessary for flowering [44]. Flowering date, flowering stem height, site conditions, and predation may affect marsh thistle seed production. In seashore meadow habitats in Tullgarn, Sweden, late-flowering marsh thistle plants produced more flower heads and generally had greater reproductive output than early-flowering plants. Reproductive output also increased with increasing height of the flowering stem (R² =0.646, P<0.0001) [48]. In a dune area north of The Hague, marsh thistle populations averaged 24.1 seed-producing flower heads/plant and produced between 300 and 2,000 seeds/plant. There were many undeveloped seeds in the flower heads; the largest percentage of undeveloped seeds occurred in populations that were inundated for parts of the summer. Shade did not impact seed development [75,76], although other research suggests that shading can limit seed production (see Shade tolerance).
Field observations made near The Hague showed that moth (Epiblema scutulana) larvae and rabbits that fed on marsh thistle stems reduced flower head production by an average of 25.2% and 31.8%, respectively [74]. When seeds were sown in dune areas near The Hague, counts 2 weeks later suggested seed predation levels of 60% to 80% [75].
Open sites are likely best for marsh thistle seedling emergence and establishment. Seedling growth and survival may be impacted by timing of germination, pollination of the parent plant, and presence of marsh thistle plant material in the soil. Growth, development, and reproductive success of plants 1 year or older are discussed in Plant development and survival.
Bare sites may favor seedling establishment. In a low-nutrient, species-rich meadow in the Czech Republic, almost no marsh thistle seedlings emerged from treatment plots where mosses, litter, and/or existing vegetation were left intact. Marsh thistle seedling emergence was greatest in plots where mosses and litter were removed [59]. In England, gaps created by domestic sheep grazing were considered important to marsh thistle seedling establishment [26].
In a spring area in central Jutland, Denmark, seedling mortality was high (85%) regardless of emergence timing, but seedlings that survived their first winter had a high probability of surviving to reproductive age [4]. Growth, reproduction, and survival of these seedlings were monitored in later life stages (see Botanical description), but spring- and fall-emerging cohorts were not studied separately [4].
Characteristics of fall- and spring-emerging seedlings in Jutland, Denmark [4] Measured (mm) or counted attribute Spring emergence Fall emergence Number of rosette leaves 2-3 2-3 Leaf width 7-11 13-23 Leaf length 40-80 90-200 Diameter of taproot 0.8-1.2 1.5-2.5 Length of taproot 20-30 20-30 Number of lateral roots 4 7-8In the Netherlands, seedlings from seeds produced by cross-pollination had significantly (P<0.05) lower overall emergence, fall emergence, and survival to 1 year old than seedlings produced by plants protected from insects. There were 100 seeds from protected plants and 100 seeds from cross-pollinated plants sown and monitored in the field [73].
Fate of cross-pollinated and noncrossed seed sown in the Netherlands [73] Seed type Cross-pollinated seed Noncrossed seed Total number of seedlings observed 44.3 57.0 Number of fall-emerging seedlings 8.7 17.2 Number of 1-year-old plants 10.5 15.8 All differences between cross-pollinated and noncrossed seeds were significant (P<0.05).Marsh thistle seedling growth may be reduced in the presence of marsh thistle leaf litter. Field observations in central Jutland, Denmark, revealed a rarity of seedlings beneath marsh thistle flowering plants or near marsh thistle rosettes. In a controlled study, marsh thistle seedling growth was monitored after 5 weeks in soils mixed with marsh thistle foliage. At a 0.25% foliage concentration, marsh thistle seedling growth was reduced by 52%, and at a concentration of 1.25%, was reduced by 65% [3].
The scientific name of marsh thistle is Cirsium palustre (L.) Scop. (Asteraceae) [17,19,30,36,54].
Hybrids:
Natural C. palustre × C. arvense hybrids occur in England and other European countries (Sledge 1975 cited in [17]). These hybrids are possible where these species grow together in North America [17].
Cirsium palustre ye una especie de planta fanerógama perteneciente a la familia Asteraceae. Ye nativa d'Europa, anque se naturalizó en nueva Zelanda y Norteamérica onde-y la considera planta invasora.
Ye una planta yerbácea añal o perenne qu'algama los 2-3 metros d'altor, con tarmos fuertes y erectos cubiertu de pequeñes escayos y tomentosos na parte cimera. Les fueyes son estreches y lobulaes y les cabezueles de la inflorescencies que se producen, nel ápiz de los tarmos en trupos recímanos, en xunu-setiembre tienen 10-15 mm de llargor coles bráctees glutinoses de color verde a púrpura. Los flósculos son de color lavanda-púrpura. Los frutos son cipseles pardes con aniellu apical y miriguanu plumosu.
Cirsium palustre describióse orixinalmente por Carlos Linneo como Carduus palustris (basónimu) en Species Plantarum, vol.2,p. 822, 1753 y tresferíu al xéneru Cirsium por Ernest Saint-Charles Cosson ex Giovanni Antonio Scopoli y espublizóse en Flora Carniolica, Editio Secunda, 2, p. 128, 1772.[1]
Cirsium: nome xenéricu que deriva de la pallabra griega: kirsos = varices ; d'esti raigañu deriva'l nome kirsion, una pallabra que paez sirvir pa identificar una planta que s'utiliza pal tratamientu d'esti tipu d'enfermedá. De kirsion, nos tiempos modernos, el botánicu francés Tournefort (1656 - 708) derivó'l nome Cirsium del xéneru.
palustre: epítetu llatín que significa "que crez nos banzaos".[2]
 Esta páxina forma parte del wikiproyeutu Botánica, un esfuerciu collaborativu col fin d'ameyorar y organizar tolos conteníos rellacionaos con esti tema. Visita la páxina d'alderique del proyeutu pa collaborar y facer entrugues o suxerencies.
Esta páxina forma parte del wikiproyeutu Botánica, un esfuerciu collaborativu col fin d'ameyorar y organizar tolos conteníos rellacionaos con esti tema. Visita la páxina d'alderique del proyeutu pa collaborar y facer entrugues o suxerencies. 
Cirsium palustre ye una especie de planta fanerógama perteneciente a la familia Asteraceae. Ye nativa d'Europa, anque se naturalizó en nueva Zelanda y Norteamérica onde-y la considera planta invasora.
Pcháč bahenní (Cirsium palustre) je hojně rozšířená dvouletá rostlina, s ostnatou lodyhou vysokou až 200 cm a nachovými květy, vyskytující se na vlhkých stanovištích.
S výjimkou jihu kontinentu roste v celé Evropě, na východě je rozšířen až za Ural, do západní i východní Sibiře. Byl zavlečen na severovýchodní i severozápadní pobřeží Severní Ameriky, do severní Afriky i na Nový Zéland. Je to rostlina vyhledávající chladnější a hlavně vlhká místa. Roste na mokrých lukách, rašeliništích, příkopech, pasekách, u pramenišť, ve světlých bažinatých lesících, na březích často rozlévajících se toků nebo jezer; limitujícím faktorem je dostatek vlhkosti. Pcháč bahenní vyžaduje hodně slunce, špatně snáší zastínění.
V České republice roste téměř po celém území, více se vyskytuje ve vyšších polohách, v teplých oblastech jižní Moravy je vzácný.[1][2][3]
Statná, hrubá, žebrovaná lodyha po celé délce silně ostnitě dorůstá v ideálních podmínkách až do výše 200 cm. Většinou je jednoduchá, někdy se větví ve stoupající větve až ve vrcholové části. Vyrůstá z kořene dlouhého 10 až 20 cm a tlustého až 2 cm s 30 až 40 tenkými postranními kořínky. Rostlina má dva druhy listů, oba jdou ostnité, na líci tmavozelené a na rubu světlezelené, jsou pokryty krátkými kadeřavými chlupy. Spodní listy vyrůstající v přízemní listové růžici jsou peřenosečné až peřenoklané, dorůstají délky 30 cm a šířky 12 cm. Střídavé lodyžní listy kopinatého tvaru jsou kratší, do 20 cm, směrem vzhůru se zmenšují, jsou peřenolaločné, úkrojky jsou nestejně trojboké, ostnitě zubaté, na lodyhu přisedají sbíhavými ostnitými křídly. Nejhornější listy lodyhy úzce kopinatého až čárkovitého tvaru jsou nedělené.
Vejčitě podlouhlé, přisedlé nebo krátce stopkaté květní úbory veliké 10 až 15 mm jsou na konci lodyhy nebo větví soustředěny do klubkatých, chudých chocholíků po 2 až 8 úborech. Zelené, na konci červenohnědé zákrovní listeny jsou štíhlé, kopinaté, na konci mají krátký osten, vnější jsou mnohem kratší než vnitřní, okraje zákrovů jsou plstnaté.
Trubkovité květy jsou většinou oboupohlavné nachové nebo zřídka bílé barvy, menší počet pouze samičích. Koruny dlouhé od 10 do 15 mm mají zvonkovitý okraj do poloviny rozeklán v pět nestejných cípů, korunní trubka má asi poloviční délku. Tyčinek je v květu pět a jsou srostlé, spodní semeník je složen ze dvou plodolistů, čnělka o délce 1,5 až 2 mm nese dvoulaločnou bliznu. Plody jsou hladké ochmýřené nažky které jsou obvejčité o velikosti asi 3 × 1 mm a mají chmýr dlouhý 8 až 12 mm se štětinkami velkými až 2 mm.[1][4][5][6]
Za optimálních podmínek se chová pcháč bahenní jako dvouletá rostlina. Brzy na jaře (koncem března), za příznivé teploty a vlhkosti, vyraší mladá rostlinka v mezeře mezi shluky rostlin ze semene z čerstvého náletu nebo ze zásoby semen v půdě. V předstihu před většinou ostatních rostlin, do 2 až 3 měsíců, vytvoří bohatou růžici přízemních listů která potlačuje okolní blízkou konkurenci. Dorostlá růžice přečká zimu a příštího roku z ní vyroste silná květná lodyha. Po dozrání semen celá rostlina usychá. Semena mohou vyklíčit ještě téhož roku po vysemenění (v říjnu), ale jejich růžice málokteré přečkají zimu, jen asi 20 %.
Leží-li semeno přes zimu v zastíněném místě, na jaře nevyklíčí. Pokud se výrazně zhorší světelné podmínky až po vyklíčení (olistěním stromu), může přečkávat rostlina ve stadiu růžice i 5 let než uhyne, případně může vyrůst nízká slabá lodyha jen s malými květy a nízkým počtem semen. Pokud se příčina stínu odstraní (ořezáním stromu) vyroste silná lodyha třeba i čtvrtým rokem. Růst lodyhy s květy je tedy podmíněn nejen dosažením určité velikosti růžice, ta vyroste i ve stínu, ale hlavně světelným stanovištěm.
Rostliny rozkvétají od června do září. Rozkvetlé květy jsou opylovány blanokřídlým a dvoukřídlým hmyzem, brouky a hlavně motýly, může u nich také dojít k samoopylení. Květy opylené vlastním pylem produkují těžší nažky které v dopadají v blízkosti mateřské rostliny a rychleji klíčí. Nažek z květů sprašených cizím pylem je mnohem více, jsou lehčí a vítr je roznáší dále do okolí, pomaleji klíčí a mají větší počet vyklíčených rostlin nedorostlých do stádia kvetení. Rostlina vyprodukuje 300 až 2000 semen, při rozptylu semen větrem většina jich nedolétne dále než 10 m.[4]
Pcháč bahenní je v Evropě považován za plevelnou rostlinu, která není příliš nebezpečná. Ponejvíce se vyskytuje na místech která nejsou příliš hospodářský využívána a dalších vhodných míst pro jeho šíření mnoho není. Pro evropské zemědělství není potenciálním ohrožením, pohlíží se na něj jako na tradiční součást evropské flory.
Jiná situace je v zemích kde byl zavlečen nedávno, hlavně v Severní Americe. Jsou tam ohromné plochy na kterých by se mohl nekontrolovaně šířit a proto mu jako invazní, potenciálně nebezpečné rostlině věnují zvláštní pozornost.[4][7]
Přízemní růžice
Lodyžní list
Rozvitý květ
Pcháč bahenní (Cirsium palustre) je hojně rozšířená dvouletá rostlina, s ostnatou lodyhou vysokou až 200 cm a nachovými květy, vyskytující se na vlhkých stanovištích.
Kærtidsel (Cirsium palustre), ofte skrevet kær-tidsel, er en 80-150 centimeter høj plante i kurvblomst-familien. Den rødbrunt anløbne stængel har tornede vinger af de nedløbende blade. De purpurrøde blomster sidder i små kurve, der er samlet i tætte klynger i spidsen af stænglen.
Kærtidsel er toårig urt, der første år danner en stor roset af fjersnitdelte blade og andet år danner en stift opret, rødbrunt anløben stængel, der kun er grenet mod spidsen. Hele palnten er stærkt tornet. De linje-lancetformede, fjerfligede blade, der ofte har en mørk rand, er nedløbende og danner derfor vinger på stænglen. De 1-1,5 centimeter brede, ustilkede kurve med purpurrøde blomster er samlet i hovedformede stande i spidsen af stænglens grene. Frugten har fjerformet fnok.
Arten er udbredt i Europa, Nordafrika og Vestasien.
I Danmark er kærtidsel temmelig almindelig i moser, på fugtige enge og i skovrydninger. Den blomstrer i juni til august.[1]
Kærtidsel (Cirsium palustre), ofte skrevet kær-tidsel, er en 80-150 centimeter høj plante i kurvblomst-familien. Den rødbrunt anløbne stængel har tornede vinger af de nedløbende blade. De purpurrøde blomster sidder i små kurve, der er samlet i tætte klynger i spidsen af stænglen.
Die Sumpf-Kratzdistel (Cirsium palustre)[1] ist eine Pflanzenart, die zur Unterfamilie der Carduoideae innerhalb der Familie der Korbblütler (Asteraceae) gehört.
Für die Region St. Gallen bei Sargans ist der Trivialname Landschnecht belegt.[2]
Die Sumpf-Kratzdistel ist eine zumeist zwei- oder mehrjährige, hapaxanthe krautige Pflanze, die Wuchshöhen von etwa 30 bis 200 (selten bis 300) Zentimetern erreicht. Ihre einzeln stehenden, aufrechten, höchstens wenig verzweigten Stängel sind über die gesamte Länge lappig oder kraus dornig geflügelt und bis zur Spitze mit stacheligen Blättern versehen. Sie fällt wegen ihrer steifen, gezackten Laubblätter auf, die oft rötlich unterlaufen, oberseits dunkelgrün, und unterseits mehr oder weniger weiß-filzig sind. Die lanzettliche bis buchtig fiederspaltige Blattspreite ist 15 bis mehr als 30 Zentimeter lang und 3 bis 10 Zentimeter breit, mit dornig gezähnten Abschnitten. Wenn Blattstiele vorhanden sind dann sind sie dornig geflügelt.
An kurzen, bis 1 Zentimeter langen Stielen stehen zwei bis acht körbchenförmige Blütenstände in Knäueln an den Stängelenden zusammen. Die etwas spinnwebig behaarten Blütenkörbe weisen eine Höhe 10 bis 20 mm und einen Durchmesser von 8 bis 13 mm auf. Die Hüllblätter stehen in fünf bis sieben Reihen. Ihre meist dunkel-purpurrot, oder selten hellrosa bis weiß gefärbten Röhrenblüten sind insgesamt 11 bis 13 mm lang mit einer 5 bis 7 mm langen Kronröhre und 3 bis 4,5 mm langen Kronzipfeln.
Die Blütezeit liegt zwischen Juli und September.
Die glänzende Achäne ist 2,5 bis 3,5 mm groß mit einem fedrig behaarten und 9 bis 11 mm langen Pappus.
Die Chromosomenzahl beträgt 2n = 34.[3]
Das natürliche Verbreitungsgebiet umfasst ganz Europa und das gemäßigte Sibirien. In den nordöstlichen Vereinigten Staaten sowie in Neufundland, Labrador, Ontario, Quebec und British Columbia in Kanada gilt sie als Neophyt.
Die Sumpf-Kratzdistel findet man vor allem in Feuchtwiesen, Sümpfen, Gräben, an Ufern oder in Auwäldern. Sie ist ein Ton- und Vernässungsanzeiger. Sie gedeiht besonders an feuchten und schattigen Standorten in Höhenlagen etwa bis zu 800 Metern. In den Allgäuer Alpen steigt sie am Osthang des Riedberger Horns in Bayern bis in eine Höhenlage von 1700 Meter auf.[4] Sie ist eine schwache Charakterart der Ordnung Molinietalia, kommt aber auch in Gesellschaften der Klassen Scheuchzerio-Caricetea oder Epilobietea angustifolii oder der Verbände Alnion oder Alno-Ulmion vor.[3]
Die Sumpf-Kratzdistel ist ein Ton- und Vernässungszeiger.[3]
Die Sumpf-Kratzdistel (Cirsium palustre) ist eine Pflanzenart, die zur Unterfamilie der Carduoideae innerhalb der Familie der Korbblütler (Asteraceae) gehört.
Für die Region St. Gallen bei Sargans ist der Trivialname Landschnecht belegt.
Błotny kagùlc (Cirsium palustre (L.) Scop.) – to je ôrt roscënë z rodzëznë astrowatëch (Asteraceae). Òn rosce m. jin. na Kaszëbach, a Kaszëbi mògą na niegò gadac kagùlc.
Cirsium palustre, the bog-thristle or Moss-thristle, is a yerbaceous biennial (or wften perennial) species o the genus Cirsium. It is a taw thistle which reaches up tae 2 metres (7 ft) in hicht.
Mýritistil (frøðiheiti Cirsium palustre) verður 50-120 cm til hæddar og veksur, sum navnið sigur, í mýrum og við áarbakkar. Henda høga, hábærsliga plantan við síni korkalittu blómu og fagra grøna leyvi hevur hugtikið mangan. Men brátt varnast tú, at hon er ikki so at koma í hóslag við, tí bæði á bløðum og leggjum vaksa hvassar tornir. Leggurin stendur steyrrættur og er harður og trækendur. Bløðini eru avlong váknvaksin við stórum flippum. Tey vaksa tøtt saman niðri við rótina og upp eftir øllum legginum. Blómurnar eru kurvavaksnar, og fleiri kurvar vaksa saman í einum tyssi ytst á leggendanum. Blómar í juli til august.
Lýsing: Um alt landið við áarbakkar og á vátlendi. Fyrsta árið bert stovnblað, annað ella triðja árið ein stinnur, strentur, langtornutur leggur. Bløðini stórflipput, niðurrend, langtornut um jaðaran. Kurvarnar korkareyðar, nógvar saman í tyssaskjóli, stuttleggjaðar; allar blómur við pípukrúnum; krúnukragin bert klovin móti miðjuni. Kurvareivbløðini takløgd og tornut.
Mýritistil (frøðiheiti Cirsium palustre) verður 50-120 cm til hæddar og veksur, sum navnið sigur, í mýrum og við áarbakkar. Henda høga, hábærsliga plantan við síni korkalittu blómu og fagra grøna leyvi hevur hugtikið mangan. Men brátt varnast tú, at hon er ikki so at koma í hóslag við, tí bæði á bløðum og leggjum vaksa hvassar tornir. Leggurin stendur steyrrættur og er harður og trækendur. Bløðini eru avlong váknvaksin við stórum flippum. Tey vaksa tøtt saman niðri við rótina og upp eftir øllum legginum. Blómurnar eru kurvavaksnar, og fleiri kurvar vaksa saman í einum tyssi ytst á leggendanum. Blómar í juli til august.
Lýsing: Um alt landið við áarbakkar og á vátlendi. Fyrsta árið bert stovnblað, annað ella triðja árið ein stinnur, strentur, langtornutur leggur. Bløðini stórflipput, niðurrend, langtornut um jaðaran. Kurvarnar korkareyðar, nógvar saman í tyssaskjóli, stuttleggjaðar; allar blómur við pípukrúnum; krúnukragin bert klovin móti miðjuni. Kurvareivbløðini takløgd og tornut.
Li tcherdon d' pré[1], c' est ene sôre di tcherdon ki vént dins les prés.
No d' l' indje e sincieus latén : Cirsium palustre
Crexhe foirt hôt, et sins waire di foyes. Li flormint est resseré ådzeu.
Corant so les prés del Hôte Fagne.
Divinltins, on l' aléve code e prétins. On l' acmoirtixheut al cujhante aiwe, pol diner ås vatches didins des påsteas et des lapaedjes.[2]
djonne
bodje, foyes et picas
flormint

 Pol discramiaedje des årtikes avou l' mot "tcherdon", loukîz cial
Pol discramiaedje des årtikes avou l' mot "tcherdon", loukîz cial Li tcherdon d' pré, c' est ene sôre di tcherdon ki vént dins les prés.
No d' l' indje e sincieus latén : Cirsium palustre
Cirsium palustre, the marsh thistle[2] or European swamp thistle, is a herbaceous biennial (or often perennial) flowering plant in the family Asteraceae.[3][4]
Cirsium palustre is a tall thistle which reaches up to 2 metres (7 ft) in height. The strong stems have few branches and are covered in small spines. In its first year the plant grows as a dense rosette, at first with narrow, entire leaves with spiny, dark purple edges; later, larger leaves are lobed. In the subsequent years the plant grows a tall, straight stem, the tip of which branches repeatedly, bearing a candelabra of dark purple flowers, 10–20 millimetres (0.4–0.8 in) with purple-tipped bracts. In the northern hemisphere these are produced from June to September. The flowers are occasionally white, in which case the purple edges to the leaves are absent.[5]
The plant provides a great deal of nectar for pollinators. It was rated first out of the top 10 for most nectar production (nectar per unit cover per year) in a UK plants survey conducted by the AgriLand project which is supported by the UK Insect Pollinators Initiative.[6]
It is native to Europe where it is particularly common on damp ground such as marshes, wet fields, moorland and beside streams. In Canada and the northern United States, it is an introduced species that has become invasive. It grows in dense thickets that can crowd out slower growing native plants.[7][5][8]
Cirsium palustre is broadly distributed throughout much of Europe and eastward to central Asia. This thistle's occurrence is linked to the spread of human agriculture from the mid-Holocene era or before.[9] It is a constant plant of several fen-meadow plant associations, including the Juncus subnodulosus-Cirsium palustre fen-meadow.[9] The flowers are visited by a wide variety of insects, featuring a generalised pollination syndrome.[10]

Cirsium palustre, the marsh thistle or European swamp thistle, is a herbaceous biennial (or often perennial) flowering plant in the family Asteraceae.
Cirsium palustre is a tall thistle which reaches up to 2 metres (7 ft) in height. The strong stems have few branches and are covered in small spines. In its first year the plant grows as a dense rosette, at first with narrow, entire leaves with spiny, dark purple edges; later, larger leaves are lobed. In the subsequent years the plant grows a tall, straight stem, the tip of which branches repeatedly, bearing a candelabra of dark purple flowers, 10–20 millimetres (0.4–0.8 in) with purple-tipped bracts. In the northern hemisphere these are produced from June to September. The flowers are occasionally white, in which case the purple edges to the leaves are absent.
The plant provides a great deal of nectar for pollinators. It was rated first out of the top 10 for most nectar production (nectar per unit cover per year) in a UK plants survey conducted by the AgriLand project which is supported by the UK Insect Pollinators Initiative.
It is native to Europe where it is particularly common on damp ground such as marshes, wet fields, moorland and beside streams. In Canada and the northern United States, it is an introduced species that has become invasive. It grows in dense thickets that can crowd out slower growing native plants.
Cirsium palustre es una especie de planta fanerógama perteneciente a la familia Asteraceae. Es nativa de Europa, aunque se ha naturalizado en nueva Zelanda y Norteamérica donde se la considera planta invasora.
Es una planta herbácea anual o perenne que alcanza los 2-3 metros de altura, con tallos fuertes y erectos cubierto de pequeñas espinas y tomentosos en la parte superior. Las hojas son estrechas y lobuladas y las cabezuelas de la inflorescencias que se producen, en el ápice de los tallos en densos racimos, en junio-septiembre tienen 10-15 mm de longitud con las brácteas glutinosas de color verde a púrpura. Los flósculos son de color lavanda-púrpura. Los frutos son cipselas pardas con anillo apical y vilano plumoso.
Cirsium palustre fue descrita originalmente por Carlos Linneo como Carduus palustris (basónimo) en Species Plantarum, vol. 2, p. 822, 1753 y transferido al género Cirsium por Ernest Saint-Charles Cosson ex Giovanni Antonio Scopoli y publicado en Flora Carniolica, Editio Secunda, 2, p. 128, 1772.[1]
Cirsium: nombre genérico que deriva de la palabra griega: kirsos = varices ; de esta raíz deriva el nombre kirsion, una palabra que parece servir para identificar una planta que se utiliza para el tratamiento de este tipo de enfermedad. De kirsion, en los tiempos modernos, el botánico francés Tournefort (1656 - 708) ha derivado el nombre Cirsium del género.
palustre: epíteto latino que significa "que crece en los pantanos".[2]

Cirsium palustre es una especie de planta fanerógama perteneciente a la familia Asteraceae. Es nativa de Europa, aunque se ha naturalizado en nueva Zelanda y Norteamérica donde se la considera planta invasora.
Suo-ohdake (Cirsium palustre) on yleinen ohdakelaji. Suo-ohdake on kaksivuotinen, joskus monivuotinen, mutta vain kerran kukkiva.
Suo-ohdake kasvaa 50–200 cm korkeaksi. Sen lehdet ovat kapealiuskaiset ja hyvin piikkiset. Varsi on punertava. Kukinnot ovat 1–2 cm leveitä mykeröitä, terälehdet purppuranpunaisia ja kehtosuomut piikkisiä.[1]
Suo-ohdake muistuttaa kyläkarhiaista, mutta sen lehdet ja piikit ovat kovemmat ja lehdet kapeammat. Suo-ohdake muistuttaa myös hieman tanakampaa piikkiohdaketta (Cirsium vulgare). Siemenillä on höyhenmäinen lentovarjo. Yksi ohdake voi tuottaa 7 000 siementä, jotka leijailevat syksyllä laajalle alalle.
Suo-ohdaketta tavataan Euroopassa ja Venäjällä. Pohjois-Amerikassa se on tulokaslaji.[2]
Suo-ohdake (Cirsium palustre) on yleinen ohdakelaji. Suo-ohdake on kaksivuotinen, joskus monivuotinen, mutta vain kerran kukkiva.
Cirsium palustre
Le cirse des marais (Cirsium palustre), également appelé bâton-du-diable, est une plante bisannuelle assez grande, poussant notamment dans les prairies humides, appartenant au genre Cirsium et à la famille des Astéracées (ou Composées).
Plante à tige érigée, ailée (ailes épineuses du bas au sommet de la tige). Longues feuilles étroites très épineuses, souvent inclinées vers le sol, devenant des lanières au sommet de la tige. capitules en grappes très denses, à fleurons pourpres. Bractées de l'involucre érigées, terminées par une pointe noire peu épineuse. Feuillage caduc.
Cirsium palustre
Le cirse des marais (Cirsium palustre), également appelé bâton-du-diable, est une plante bisannuelle assez grande, poussant notamment dans les prairies humides, appartenant au genre Cirsium et à la famille des Astéracées (ou Composées).
Il cardo di palude (nome scientifico Cirsium palustre (L.) Scop., 1772) è un'alta pianta erbacea angiosperma dicotiledone a ciclo biennale, molto spinosa, appartenente alla famiglia delle Asteraceae.[1][2]
Il nome del genere (Cirsium) deriva dalla parola greca kirsos = varice; da questa radice deriva poi la denominazione Kirsion, un vocabolo che sembra servisse ad identificare una pianta usata per curare questo tipo di malattia. Da kirsion in tempi moderni il botanico francese Tournefort (1656 - 708) derivò il nome Cirsium dell'attuale genere.[3][4]
Il nome italiano “Cardo” è abbastanza generico in quanto nel linguaggio comune si riferisce a diversi generi e specie di piante. Tra i generi che vengono chiamati direttamente “cardo”, oppure hanno una o più specie che comunemente si chiamano con questo nome citiamo: Carduus, Carduncellus, Carlina, Centaurea, Cnicus, Cynara, Echinops, Galactites, Jurinea, Onopordum, Scolymus, Silybum, Tyrimnus, tutti della famiglia delle Asteraceae. Ma anche in altre famiglie abbiamo dei generi con delle specie che volgarmente vengono chiamate “cardi” : il genere Eryngium della famiglia delle Apiaceae o il genere Dipsacus della famiglia delle Dipsacaceae.
Il binomio scientifico della pianta di questa voce è stato proposto inizialmente da Carl von Linné (1707 – 1778) biologo e scrittore svedese, considerato il padre della moderna classificazione scientifica degli organismi viventi, perfezionato successivamente dal medico e naturalista italiano Giovanni Antonio Scopoli (Cavalese, 3 giugno 1723 – Pavia, 8 maggio 1788) nella pubblicazione ”Flora Carniolica 2” del 1772.[5]
L'epiteto specifico (palustre = delle paludi) deriva da uno degli habitat tipici di questo fiore.[6]
Le piante possono arrivare ad un'altezza di 1,5 – 2 metri (minimo 50 cm – in qualche caso sono stati rilevati individui alti fino a 3 metri[7]). La forma biologica della specie è emicriptofita bienni ("H bienn"); ossia sono piante a ciclo riproduttivo biennale che si riproducono per mezzo di gemme poste al suolo. Nel corso del primo anno presentano solamente una rosetta fogliare mentre nel secondo anno fioriscono completamente ma una sola volta (specie monocarpica).[7] L'indumento in generale è molto spinoso e peloso per peli di tipo tricomi articolati.[8][9][10][11][12][13][14]
Le radici consistono in ammassi di filamenti radicali fibrosi.
Il fusto normalmente è eretto, singolo e ramoso solo nella parte alta ed è alato per quasi tutta la sua lunghezza (in quanto i margini delle foglie sono decorrenti e pungenti). Può presentarsi lievemente ricurvo all'apice a causa del peso dei fiori che normalmente si addensano in quella parte.


Le foglie sono oblunghe (o strettamente ellittiche o anche oblanceolate) e sinuato-pennatopartite (segmenti separati da grandi seni) con lobi triangolari terminanti in una spina. Il margine delle foglie è dentato – pungente; sono inoltre sessili con base ristretta. Sulla pagine superiore (adassiale) sono verdi e ispide (o anche glabre); su quella inferiore (abassiale) sono bianco – tomentose/ragnatelose (soprattutto i primi getti) con tricomi articolati.
Dimensioni delle foglie (quelle basali) : larghezza 2 – 4 cm; lunghezza da 12 a 20 cm (massimo 10 cm x 30 cm). Le spine possono essere lunghe fino a 4 – 6 mm. L'area centrale indivisa è larga 5 – 8 mm.




L'infiorescenza consiste in capolini da sessili a più o meno peduncolati (da 0 a 1 cm) a forma di pannocchia ombrelliforme (riuniti in densi grappoli di 2 - 8 capolini). Il capolino può essere lievemente tomentoso. Lunghezza del peduncolo da 0 a 1 cm. Dimensione dei capolini: larghezza 8 mm; lunghezza 15 mm. I fiori sono racchiusi in un involucro a forma di bicchiere (o a forma campanulata) lungo 1 - 1,5 cm; diametro 0.7 – 1,1 cm e circondato da brattee o squame percorse da sottili ragnatele tomentose con tricomi non ramificati. Le squame dell'involucro sono embricate (in 5 – 7 serie) e chiazzate di brunastro, mentre hanno l'apice pungente di colore nerastro; la forma è di tipo da lanceolato-lineare (quelle interne) a ovata (quelle esterne). Dimensioni delle squame inferiori 1,2 per 4 mm; quelle superiori 1 per 10 – 12 mm. All'interno dell'involucro un ricettacolo fa da base ai fiori.
I fiori sono ermafroditi e tutti di forma tubulosa (il tipo ligulato, come nella maggioranza delle Asteraceae, è assente). Sono inoltre tetraciclici (calice – corolla – androceo – gineceo) e pentameri (ogni verticillo ha 5 elementi).
 , [C (5), A (5)], G 2 (infero), achenio[15]
, [C (5), A (5)], G 2 (infero), achenio[15]
Il frutto è un achenio cilindrico con pappo piumoso formato da molte file di peli barbosi riuniti alla base. Il pappo ha la funzione di aiutare la dispersione del seme portato quindi dal vento. Essendo la pianta anche abbastanza alta il successo con questo tipo di dispersione è molto rilevante. Dimensione del pappo 10 – 13 mm. Dimensione dell'achenio: 2,5 – 3,5 mm.
Dal punto di vista fitosociologico alpino la specie di questa voce appartiene alla seguente comunità vegetale[18]:
Per l'areale completo italiano la specie di questa voce appartiene alla seguente comunità vegetale:[19]
Descrizione. L'alleanza Molinion caeruleae è relativa ai prati umidi non concimati frequentati soprattutto dalla specie Molinia caerulea. Questa cenosi si sviluppa in prevalenza su suoli da subacidi a neutro-alcalini, talora torbosi e soggetti a parziale inaridimento estivo. Distribuzione dell'alleanza: dall'Italia settentrionale fino in Calabria. L’alleanza è ampiamente distribuita nell’Europea temperata. Queste comunità sono delle praterie seminaturali che, in assenza di sfalcio, evolvono, in tempi anche brevi, in comunità legnose dominate dalle specie di betulle e faggete.
Specie presenti nell'associazione: Cirsium palustre, Molinia caerulea, Serratula tinctoria, Equisetum palustre, Potentilla erecta, Succisa pratensis, Valeriana dioica, Sanguisorba officinalis, Genista tinctoria, Valeriana officinalis, Carex pallescens, Carex panicea, Carex davalliana, Schoenus nigricans, Lathyrus pratensis, Festuca pratensis, Festuca nigrescens, Parnassia palustris, Briza media, Holcus lanatus, Juncus articulatus, Selinum carvifolia, Potentilla erecta, Stachys officinalis, Trifolium pratense, Trifolium dubium, Trifolium patens, Centaurea nigrescens, Ranunculus acris e Rumex acetosa.
Altre alleanze per questa specie sono:[19]
La famiglia di appartenenza di questa voce (Asteraceae o Compositae, nomen conservandum) probabilmente originaria del Sud America, è la più numerosa del mondo vegetale, comprende oltre 23.000 specie distribuite su 1.535 generi[20], oppure 22.750 specie e 1.530 generi secondo altre fonti[21] (una delle checklist più aggiornata elenca fino a 1.679 generi)[22]. La famiglia attualmente (2021) è divisa in 16 sottofamiglie.[1][11][23]
Cardueae è una delle 4 tribù della sottofamiglia Carduoideae. La tribù Cardueae a sua volta è suddivisa in 12 sottotribù (la sottotribù Carduinae è una di queste). Il genere Cirsium elenca 435 specie con una distribuzione cosmopolita, 35 delle quali sono presenti spontaneamente sul territorio italiano.[2][11][12][13][24][25]
Il genere di questa voce è inserito nel gruppo tassonomico della sottotribù Carduinae.[13] In precedenza provvisoriamente era inserito nel gruppo tassonomico informale "Carduus-Cirsium Group".[11] La posizione filogenetica di questo gruppo nell'ambito della sottotribù è abbastanza vicina al "core" della sottotribù (con il genere Carduus forma un "gruppo fratello") e dalle analisi molecolari è stato calcolato in 7,2 milioni di anni fa la separazione di questo genere dal resto del gruppo (è stato l'ultimo a separarsi).[24][25]
Il genere Cirsium spesso viene botanicamente “confuso” con altri generi come quello del Carduus o Cnicus (e di altri ancora). Le specie del primo genere ad esempio sono molto simili a quelle del Cirsium, anche se una certa distinzione è possibile servendosi dell'aspetto del pappo (in Cirsium è formato da setole piumose; mentre in Carduus è composto da pagliette denticolate scabre).[26]
Il genere Cirsium appartiene alla tribù delle Cardueae (da alcuni autori indicata come Cynareae), tribù che il Sistema Cronquist assegna alla sottofamiglia Cichorioideae e che invece la classificazione APG colloca nella sottofamiglia Carduoideae.[27].
Il numero cromosomico di Cirsium palustre è 2n=34.[14][28]
Il basionimo per questa specie è: Carduus palustris L, 1753.[18]
I caratteri distintivi di questa specie nell'ambito del genere sono:[29][30]
Questi caratteri sono condivisi con le seguenti specie (sono indicati anche alcuni caratteri distintivi per ogni specie):[30]
Sandro Pignatti nella sua ”Flora d'Italia” (1982) segnala una varietà presente in Italia: Cirsium palustre var. horridum Pospichal (le spine sono più robuste, fino a 13 mm di lunghezza; oppure i fiori hanno le corolle più o meno decolorate). Varietà che però non è più stata confermata in seguito o inserita nelle più recenti checklist della flora spontanea italiana; e attualmente viene considerato un sinonimo.[31] In alcune checklist nell'ambito della tassonomia di questa voce viene inclusa anche la specie Cirsium bourgaeanum Willk. [32]
Nell'elenco seguente sono indicati alcuni ibridi intraspecifici:[33]
La specie “Cardo di palude”, in altri testi, può essere chiamata con nomi diversi. L'elenco seguente indica alcuni tra i sinonimi più frequenti:[31]
Le foglie e i fiori hanno un certo interesse alimentare: specialmente i giovani germogli cotti. Ma anche i gambi (opportunamente pelati) si possono mangiare come gli asparagi.[34]
Dai semi della pianta si può ricavare dell'olio.[34]
È una pianta infestante (specialmente nei pascoli dell'America del Nord) dove a volte crea estese aree impenetrabili. Sembra che il potenziale di diffusione sia accresciuto notevolmente dalla particolari condizioni ambientali. In queste zone, per ridurre l'infestazione, si usano vari metodi sia meccanici che chimici (taglio selettivo in prossimità del suolo o taglio dei fiori prima della dispersione dei semi; oppure tramite opportuni erbicidi).[7]
Il Cirsio di plaude in altre lingue viene chiamata nei seguenti modi:




Rosetta basale
Rosetta del primo anno
Il cardo di palude (nome scientifico Cirsium palustre (L.) Scop., 1772) è un'alta pianta erbacea angiosperma dicotiledone a ciclo biennale, molto spinosa, appartenente alla famiglia delle Asteraceae.
De kale jonker (Cirsium palustre) is een twee- of meerjarige vederdistel in de onderfamilie Carduoideae van de composietenfamilie. Ze overwintert met winterknoppen onder of net boven de grond, ze is monocarp.
De 60-120 cm hoge plant is zeer algemeen op open, vochtige omgewerkte grond. Langs de stengel staan de stekelige, tot aan de top gevleugelde bladeren. Deze bladeren zijn bochtig veerspletig, met 2 tot 3 lobbige slippen. De stengel is meestal hooguit aan de top vertakt.
De roodpaarse bloemhoofdjes zijn kort gesteeld en staan in kluwens. Zeldzaam komen ook witte bloemhoofdjes voor. De kluwens zijn vrijwel bladloos. De bloeiperiode valt in de periode juni t/m september. De omwindselbladen zijn vaak paars aangelopen en zwak stekelig.
De kruising Cirsium dissectum × palustre is een tussenvorm tussen de beide ouders die op dezelfde plaatsen als Cirsium dissectum voorkomt.
Het verspreidingsgebied beslaat Noord- en Midden-Europa, en loopt in Azië door tot in West-Siberië. In Midden-Siberië is ze zeldzaam. In Noord-Amerika is ze ingevoerd.
Ze komt o.a. voor in de plantensociologische groep Molinietalia. Ze is gastheerplant voor Aethes cnicana, Epiblema cirsiana, Scrobipalpa acuminatella, Scrobipalpa pauperella en in Finland voor Autographa gamma.

De kale jonker (Cirsium palustre) is een twee- of meerjarige vederdistel in de onderfamilie Carduoideae van de composietenfamilie. Ze overwintert met winterknoppen onder of net boven de grond, ze is monocarp.
Fries: Houtstikel Engels: Marsh thistle Frans: Cirse des marais Duits: Sumpf-KratzdistelMyrtistel er ein fleirårig art i korgplantefamilien. Planta er utbreidd over Europa, Asia og Nord-Afrika. Myrtistel har rak stengel med korte greiner øvst, oftast litt raudbrun. Han har buktfinna mørkgrøne blad med sylspisse broddar og mørkfiolette korgdekkblad utan kvass odd. Planten blomstrar kring juli månad på grasmyr og våt eng. Han veks i Noreg nord til Troms, og er funnen til fjells opp til 1000 m.
Myrtistel er ein fleirårig art i korgplantefamilien. Planta er utbreidd over Europa, Asia og Nord-Afrika. Myrtistel har rak stengel med korte greiner øvst, oftast litt raudbrun. Han har buktfinna mørkgrøne blad med sylspisse broddar og mørkfiolette korgdekkblad utan kvass odd. Planten blomstrar kring juli månad på grasmyr og våt eng. Han veks i Noreg nord til Troms, og er funnen til fjells opp til 1000 m.
Ostrożeń błotny (Cirsium palustre (L.) Scop.) – gatunek rośliny należący do rodziny astrowatych. Występuje w całej Europie oraz na Syberii Zachodniej i Wschodniej, rozprzestrzenił się także gdzieniegdzie poza tymi obszarami swojego rodzimego występowania[2]. W Polsce gatunek pospolity, zarówno na niżu, jak i w niższych położeniach górskich.
Kwiatostan
Łodyga
Liść
Roślina dwuletnia. W pierwszym roku wegetacji tworzy różyczkę liści, w drugim łodygę z kwiatami i owocami. Roślina kwitnie od lipca do września, jest owadopylna.
Rośnie na torfowiskach, wilgotnych łąkach, w zaroślach. W górach sięga po regiel dolny. W uprawach rolnych, a szczególnie na łąkach jest uciążliwym chwastem. Hemikryptofit. W klasyfikacji zbiorowisk roślinnych gatunek charakterystyczny dla O. Molinieatalia[3].
Tworzy mieszańce z większością gatunków ostrożeni, przeważnie z ostrożeniem łąkowym i ostrożeniem warzywnym[4].
Ostrożeń błotny (Cirsium palustre (L.) Scop.) – gatunek rośliny należący do rodziny astrowatych. Występuje w całej Europie oraz na Syberii Zachodniej i Wschodniej, rozprzestrzenił się także gdzieniegdzie poza tymi obszarami swojego rodzimego występowania. W Polsce gatunek pospolity, zarówno na niżu, jak i w niższych położeniach górskich.
Cirsium palustre é uma espécie de planta com flor pertencente à família Asteraceae.
A autoridade científica da espécie é (L.) Scop., tendo sido publicada em Flora Carniolica, Editio Secunda 2: 128. 1772.[1]
O seu nome comum é cardo-palustre.[2]
Trata-se de uma espécie presente no território português, nomeadamente em Portugal Continental.
Em termos de naturalidade é nativa da região atrás indicada.
Não se encontra protegida por legislação portuguesa ou da Comunidade Europeia.

Cirsium palustre é uma espécie de planta com flor pertencente à família Asteraceae.
A autoridade científica da espécie é (L.) Scop., tendo sido publicada em Flora Carniolica, Editio Secunda 2: 128. 1772.
O seu nome comum é cardo-palustre.
Močvirski osat (znanstveno ime Cirsium palustre) je trajnica iz družine nebinovk.
Močvirski osat zraste do 2 metra visoko. Listi so na spodnji strani dlakavi, spodnji se iztezajo po steblu. Steblo se proti vrhu razdeli, na vrhu vsake veje pa se razvije po več cvetnih koškov. Koški so majhni, v premeru merijo do 15 mm, ovojkovi listi pa se končujejo s trnom.Cvetovi so rdečevijolične barve, redkeje so lahko tudi beli. Najpogosteje to vrsto osata najdemo na mokrih travnikih, gozdnih močvirjih in vlažnih posekah od severne Evrope do osrednje Azije. Kasneje so ta osat zanesli tudi v Severno Ameriko, kjer je postal invazivna vrsta.
Močvirski osat (znanstveno ime Cirsium palustre) je trajnica iz družine nebinovk.
Kärrtistel (Cirsium palustre) är en art tillhörande familjen korgblommiga växter. Den förekommer i Sverige både som två- och flerårig.
Arten blir mellan 50 och 250 centimeter hög. Den har smala mörkgröna parflikiga blad som har taggig kant och blekgrön undersida. Den blommar mellan juni och september, men i Sverige börjar blomningen normalt inte förrän i juli. Kärrtistel får då 2 till 8 stycken blomkorgar som är 7 till 10 millimeter breda och mörklila till färgen. Artens frö är försett med fjäderpensel. Hela plantan kan ofta ha en brunrödaktig ton. Holkfjällen är glest håriga och ibland klibbiga. Kärrtistel växer ofta, som namnet antyder, vid kärr men även längs diken och på fuktiga betesmarker och där det finns öppen fuktig mull- och torvmark. Bildar hybrider tillsammans med brudborste, jordtistel och kåltistel. Artepitetet palustre kommer ifrån latinets palus som betyder kärr, och syftar på den typiska biotopen.
Kärrtistel är mycket vanlig i hela Norden, med undantag för fjälltrakterna och Island.
Kärrtistel (Cirsium palustre) är en art tillhörande familjen korgblommiga växter. Den förekommer i Sverige både som två- och flerårig.
Arten blir mellan 50 och 250 centimeter hög. Den har smala mörkgröna parflikiga blad som har taggig kant och blekgrön undersida. Den blommar mellan juni och september, men i Sverige börjar blomningen normalt inte förrän i juli. Kärrtistel får då 2 till 8 stycken blomkorgar som är 7 till 10 millimeter breda och mörklila till färgen. Artens frö är försett med fjäderpensel. Hela plantan kan ofta ha en brunrödaktig ton. Holkfjällen är glest håriga och ibland klibbiga. Kärrtistel växer ofta, som namnet antyder, vid kärr men även längs diken och på fuktiga betesmarker och där det finns öppen fuktig mull- och torvmark. Bildar hybrider tillsammans med brudborste, jordtistel och kåltistel. Artepitetet palustre kommer ifrån latinets palus som betyder kärr, och syftar på den typiska biotopen.
Kärrtistel är mycket vanlig i hela Norden, med undantag för fjälltrakterna och Island.
Дворічна або однократно квітуча багаторічна трава заввишки 50–150(300) см. Коріння мочкувате. Стебла від нерозгалужених до розгалужених на верхівці, крилаті, колючі, рифлені, шерстисті, зазвичай темно-червонуваті. Листки перистороздільні, з 2–3-лопатевими частками, паутинисто-повстяні, на краях із короткими шипуватими віями; лопаті й зубці листя і крила стебел закінчуються тонкими буро-світло-жовтими колючками 1–5 мм довжиною. Обгортка1 яйцеподібна, досить густо-паутиниста, буро-пурпурова. 1–1.5 см квіткові голови зібрані по від мало до багато у густих скупченнях. Квітки пурпурні (іноді білі або рожеві), відгин віночка менше ніж до середини 5-роздільний. Папуси 9–11 мм. Сім'янкоподібні плоди від рудувато-коричневого до солом'яного забарвлення, 2.5–3.5 мм з комірами 0.1–0.2 мм, блискучі. Квіти відвідує широкий спектр комах.
Азія: Казахстан, Росія; Європа: Білорусь, Естонія, Латвія, Литва, Молдова, Україна, Австрія, Бельгія, Чехія, Німеччина, Угорщина, Нідерланди, Польща, Словаччина, Швейцарія, Данія, Фарерські острови, Фінляндія, Ірландія, Норвегія, Швеція, Велика Британія, Албанія, Боснія і Герцеговина, Хорватія, Італія, Чорногорія, Румунія, Сербія, Словенія, Франція, Португалія, Іспанія. Введено: Португалія — Азорські острови, Нова Зеландія, Канада, Сент-П'єр і Мікелон, США.
Населяє болота, вологі ліси, луки, пасовища, канави, береги, ріллю, парові землі, скельні басейни. Індикатор вологих і глинистих земель.
В Україні зростає на болотах і заболочених луках — на б. ч. території, крім Степу і Криму[1].
_Scopoli.JPG)


1 сукупність верхніх листків або приквітків біля основи суцвіття
Cirsium palustre là một loài thực vật có hoa trong họ Cúc. Loài này được (L.) Coss. ex Scop. mô tả khoa học đầu tiên năm 1772.[1]
Cirsium palustre là một loài thực vật có hoa trong họ Cúc. Loài này được (L.) Coss. ex Scop. mô tả khoa học đầu tiên năm 1772.
Cirsium palustre (L.) Scop., 1772
Бодя́к боло́тный (лат. Círsium palústre) — травянистое растение, вид рода Бодяк семейства Сложноцветные (Asteraceae).
Двулетник с колючими листьями и крылатым колючим стеблем, со скученными на верхушке стебля корзинками. Часто в окраске листьев и стеблей имеется фиолетовый оттенок.
Двулетнее растение (редко — монокарпический многолетник) 50—150(200) см высотой. Стебель прямостоячий, узко- и колючекрылатый, с паутинистым опушением, светло-зелёный, часто с буровато-фиолетовым оттенком, в верхней половине иногда слабо разветвлённый.
Листья 10—20 см длиной и 5—10 см шириной, сверху жёлто-зелёные, часто с буровато-фиолетовым оттенком, снизу более светлые, на коротких крылатых черешках. Пластинка обратноланцетная до продолговато-ланцетной в очертании, перисто-рассечённая на ланцетные или продолговато-ланцетные сегменты с волнистым зубчатым краем, с неравными шипами на концах зубцов. На верхней стороне пластинки опушение рассеянное, снизу — густое, паутинистое.
Корзинки многочисленные, скученные на конце стебля или ветвей, 10—20 мм в поперечнике. Обёртка многорядная, листочки её 3—13×1,5—2 мм, жёлто-коричневые, по крайней мере ближе к верхушке часто с фиолетовым оттенком, ланцетной формы, с редким паутинистым опушением, на конце с шипиком. Внутренние листочки обёртки с нитевидным плёнчатым придатком. Все цветки трубчатые, венчик 10—15 мм длиной, пятилопастный, сиреневый, редко белый. Цветоложе плоское, с густыми щетинками.
Семянки обратноконической формы, 2,5—4 мм длиной и 1,2—1,4 мм шириной, на верхушке с беловатым хохолком 8—12 мм длиной. Поверхность матовая, беловатая, желтоватая или с пурпурным оттенком, неясно бороздчатая.
Евросибирское растение, встречающееся по сырым лугам и пастбищам, на пересыхающих болотах, в заболоченных лесах, в долинах рек, у канав.
Бодяк занесён на северо-восток Северной Америки (впервые обнаружен в 1902 году), где является инвазивным видом, быстро распространяющимся по влажным местообитаниям.
Диагноз вида был опубликован Карлом Линнеем в книге Species plantarum: Carduus foliis dentatis decurrentibus: margine spinosis, floribus racemosis erectis, pedunculis inermibus — «Carduus с зубчатыми низбегающими листьями, по краю колючими, с цветками в прямостоячих кистях, с цветоносами без колючек».
Номенклатурные:
Таксономические:
Бодя́к боло́тный (лат. Círsium palústre) — травянистое растение, вид рода Бодяк семейства Сложноцветные (Asteraceae).
Двулетник с колючими листьями и крылатым колючим стеблем, со скученными на верхушке стебля корзинками. Часто в окраске листьев и стеблей имеется фиолетовый оттенок.



_Scopoli.JPG)

_Scopoli.JPG)

















_Scopoli.JPG)


{kind=link}
{kind=link}